1 abr 2011
TEMA 9. OLFATO
9. OLFATO
El olfato u olfacción es el sentido encargado de detectar y procesar los olores. Es un quimiorreceptor en el que actúan como estimulante las partículas aromáticas u odoríferas desprendidas de los cuerpos volátiles, que ingresan por el epitelio olfatorio ubicado en la nariz, y son procesadas por el sistema olfativo.
La nariz humana distingue entre más de 10.000 aromas diferentes. El olfato es el sentido más fuerte al nacer.

Las sustancias odorantes son compuestos químicos volátiles transportados por el aire. Los objetos olorosos liberan a la atmósfera pequeñas moléculas que percibimos al inspirar. Estas moléculas alcanzan la mucosa olfativa, que consta de tres tipos característicos de células: las células olfativas sensoriales, las células de sostén y las células basales, que se dividen aproximadamente una vez al mes y reemplazan a las células olfativas moribundas. Los 20 o 30 millones de células olfativas humanas contienen, en su extremo anterior, una pequeña cabeza con cerca de 20 pequeños filamentos sensoriales (cilios). El moco nasal acuoso transporta las moléculas aromáticas a los cilios con ayuda de proteínas fijadoras; los cilios transforman las señales químicas de los distintos aromas en respuestas eléctricas.

Las prolongaciones nerviosas de las células olfativas alcanzan el bulbo olfatorio a través de micro-orificios del cráneo; el bulbo es una porción anterior del cerebro, que se ocupa de la percepción de los olores. Estas prolongaciones nerviosas terminan en los glomérulos, pequeñas terminaciones de celulas olfativas de forma esférica donde se procesan las señales aromáticas que luego son conducidas por células receptoras especiales. La información llega primero al sistema límbico y al hipotálamo, regiones cerebrales ontogenéticamente muy antiguas; responsables de las emociones, sentimientos, instintos e impulsos, tales regiones almacenan también los contenidos de la memoria y regulan la liberación de hormonas. Por este motivo, los olores pueden modificar directamente nuestro comportamiento y las funciones corporales. Sólo más tarde parte de la información olorosa alcanza la corteza cerebral y se torna consciente.

{kind=link}
9.1 QUÍMICA DE LOS OLORES
Estructura histológica de la Mucosa Olfatoria
La mucosa olfatoria es una zona amarilla de la mucosa nasal, que está constituida por el epitelio olfatorio que contiene al quimiorreceptor, y por la lámina propia. En el ser humana, como también en los demás animales microsmáticos, el epitelio olfatorio ocupa una pequeña superficie; cubre aproximadamente 5cm2 del receso posterior dorsal de cada fosa nasal. Bajo el epitelio olfatorio subyace una delgada lámina propia conjuntiva, en la cual se encuentran las glándulas de Bowman, los axones olfatorios que conforman la fila olfatoria y vasos sanguineos.
- Neuroepitelio: En él se localizan:
v Células de soporte: Ricas en enzimas que metabolizan a las sustancias odoríferas. Ellas poseen inclusiones que corresponden a uun pigmento pardusco, semejante a la lipofuscina y responsable del color amarillo de la mucosa.
v Células basales: Reservorio de neuronas, se consideran células progenitoras indiferenciadas.
v Células microvellosas: Su estructura fina y similar a las neuronas, permite considerarlas como quimiorreceptores secundarios.
v Neuronas olfatorias: Son células bipolares. Sus dendritas se proyectan hacia la cavidad nasal, finalizando en un engrosamiento con cilios quimiosensores (primera superficie de contacto). Sus axones son amielínicos y forman el nervio olfatorio que atraviesa la lámina cribosa del hueso etmoides, llegando al bulbo olfatorio donde hacen sinapsis principalmente con las células mitrales y también con las células en penacho.

- Lámina propia o submucosa: El epitelio olfatorio se ubica sobre una lámina propia muy celular, conformada por un tejido conjuntivo laxo con un vasto plexo de capilares sanguíneos; se observan en ella células tales como macrófagos, células plasmáticas y células pigmentarias, linfocitos B y células cebadas mucosales. Además, contiene las glándulas de Bowman, haces de axones olfatorios (fila olfatoria) y plexos venosos y linfáticos profundos. Este tejido conjuntivo laxo se continua con el tejido conjuntivo denso del periostio de la lámina cribiforme.
Sobre el neuroepitelio se localiza la capa de moco producida por las glándulas de Bowman. Este moco está constituido por:
Ø Agua.
Ø Mucopolisacáridos
Ø Proteínas.
Ø Enzimas.
Ø OBP (Proteínas de unión a fragancias).
Ø Olfatomedinas (Factores neurotróficos que favorecen la transformación
Ø de las células basales).
Típicamente, el olor de cualquier material comprende una mezcla compleja de multitud de moléculas olorosas de distintos tipos, cada una en una concentración determinada, aunque en ocasiones predomina el efecto de un elemento puro, compuesto de un único tipo de moléculas olorosas que determina el olor dominante.
Un intento inicial de abordar el problema de estudio de los olores desde el punto de vista químico fue tratar de establecer relaciones entre la estructura molecular y las propiedades olorosas. Sin embargo, esta relación no es en absoluto unívoca y hay muchos más factores a tener en cuenta (si bien existen patrones comunes a muchas moléculas olorosas).
Las moléculas que pueden ser detectadas por nuestro sentido del olfato deben tener ciertas propiedades, como por ejemplo ser lo suficientemente pequeñas como para ser volátiles (masa molecular relativa mayor de 30 a 300 g·mol-1), de modo que puedan vaporizarse, alcanzar la nariz y luego disolverse en la mucosa olfativa. Otros factores importantes son las interacciones internas (moléculas polares o no polares), la distribución de átomos, la distribución de carga, y la posibilidad de que se produzcan rotaciones estructurales. Finalmente, las relaciones (fuerzas) intermoleculares también tienen mucha importancia, y determinan finalmente su volatilidad y solubilidad en agua, entre otros factores.

Algunos de los compuestos olorosos más destacados tienen estructuras en anillo con electrones no localizados, lo que les da una especial estabilidad desde el punto de vista químico. Los químicos llaman a esta propiedad “aromaticidad”, porque los primeros compuestos que se descubrieron con dicha estructura presentaban olor, aunque muchas (pero no todas) de estas moléculas aromáticas (en el sentido químico) tienen de hecho olor.
Los receptores químicos del olfato son:
- La glándula pituitaria roja: Se ubica en la parte inferior de la fosa nasal y está recubierto por numerosos vasos sanguíneos que calientan el aire.
- La glándula pituitaria amarilla: Se ubica en la parte superior de las fosas nasales y presenta tres capas:
- Células de sostén
- Células olfatorias
- Células basales
Las células olfatorias son células nerviosas receptoras de estímulos químicos provocados por los vapores. En la pituitaria amarilla se encuentran las glándulas mucosas de Bowman, que libera un líquido que mantiene húmedo y limpio el epitelio olfatorio.

Para estimular éstas es necesario que las sustancias sean volátiles, es decir, han de desprender vapores que puedan penetrar en las fosas nasales, y que sean solubles en agua para que se disuelvan en el moco y lleguen a las células olfatorias. Éstas transmiten un impulso nervioso al bulbo olfatorio y de este a los centros olfatorios de la corteza cerebral, que es donde se aprecia e interpreta la sensación de olor. Se cree que existen 7 tipos de células olfatorias, cada una de las cuales sólo es capaz de detectar un tipo de moléculas, éstas son:
- Alcanforado: olor a naftalina.
- Almizclado: olor a almizcle.
- Mentolado.
- Etéreo: olor a fluidos de limpieza en seco.
- Picante.
- Pútrido.
En el año 1991 se descubrieron los primeros genes de las proteínas receptoras del olor. Estas moléculas receptoras residen en la membrana de células sensoriales, que retienen un aroma y envían el mensaje correspondiente al cerebro a través de una cadena de reacciones químicas. En 1996 fue caracterizado el primer receptor olfativo humano.
El olor dulce existe, al igual que los olores amargos. Se asemejan tanto como con los gustos
9.2 TRANSDUCCIÓN OLFATIVAINCIPIOS de Transducción
Principios comunes:
1. Transduccion = conversión de un estímulo en un cambio del potencial de membrana
2. Amplificación de la señal, con bajo nivel de ruido
3. Adaptación a fuertes, prolongados o repetidos estímulos
4. Integración de señales
Transducción olfativa
En el sistema olfatorio la transducción significa la conversión de señales químicas en una señal eléctrica que puede transmitirse al sistema nervioso central. Los pasos en la transducción olfatoria sin los siguientes:
1. Moléculas odorantes se unen a receptores sobre los cilios de las células olfatorias.
2. Estos receptores esta acoplados a adenilciclasa a través de una proteína G. Cuando la sustancia odorante esta unida a la proteína G, esta se activa y esto activa a su vez a una adenilcilasa.
3. La adenilciclasa cataliza la conversión de ATP en AMPc. La concentración intracelular de AMPc aumenta y abre canales de Na+ en la membrana celular del receptor olfatorio. Cuando los canales de Na+ se abren, la membrana de las células receptoras se despolarizan. Este potencial despolarizante de receptor aproxima el potencial de membrana al umbral y despolariza el segmento inicial del axón del nervio olfatorio. A continuación se generan potenciales de acción que se propagan a lo largo de los axones del nervio olfatorio en dirección del bulbo olfatorio.

Codificación del impulso olfatorio
No se sabe con exactitud de que manera se codifican los estímulos olfatorios. Se conoce la siguiente información:
1. Las células receptoras olfatorias reaccionan a varios odorantes.
2. Las células receptoras olfatorias muestran diferentes respuestas a los mismos odorantes.
3. Cuando se examina la respuesta a un determinado odorante en muchos receptores, surgen diferentes patrones para diversos odorantes.
Cada odorante produce un patrón único de actividad dentro de una población de receptores y dicho patrón se transmite al sistema nervioso central donde es interpretado como ese odorante.
Vías olfatorias
Los axones de las células receptoras abandonan el epitelio olfatorio, pasan a través de la lámina cribosa y hacen sinapsis sobre dendritas apicales de células mitrales en el bulbo olfatorio. Esta sinapsis tiene lugar en grupos a los que se denomina glomérulos. En los glomérulos convergen casi 1000 axones de receptores olfatorios sobre una célula mitral.
Las células mitrales se disponen en una sola capa dentro del bulbo olfatorio y poseen dendritas laterales y apicales. El bulbo olfatorio también contiene células granulosas y periglomerulares. Las células granulosas y perigromerulares son interneuronas inhibitorias que hacen sinapsis dendro-dendriticas sobre las células mitrales vecinas. Los impulsos inhibitorios pueden generar inhibición lateral que puede agudizar la información enviada al sistema nervioso central. Las células mitrales del bulbo olfatorio se prolongan hacia los centros elevados del sistema nervioso central. Conforme el fascículo olfatorio se aproxima a la base del cerebro, se divide en dos fascículos principales, un fascículo lateral que establece sinapsis en la corteza olfatoria primaria que incluye la corteza piriforme, y el fascículo olfatorio medial que se prolonga hasta la comisura anterior y el bulbo olfatorio contralateral.
Bibliografía
http://insn.die.upm.es/docs/INSN0506-ElOlfatoYElGusto-JMG-v18-IncluyeInsercionRepaso.pdf
bosquejodemedico.files.wordpress.com/.../alteraciones-del-olfato.doc
es.wikipedia.org/wiki/Olfato
TEMA 8. GUSTO
8. GUSTO
¢ Localizado en la cavidad bucal, específicamente en la lengua.
¢ Forma parte de uno de los órganos especializados de los sentidos.
¢ Su función es percibir los sabores de sustancias químicas disueltas en saliva.
¢ La superficie de la lengua esta cubierta por membranas gustativas que El gusto es uno de los sentidos, consiste en registrar el sabor e identificar determinadas sustancias solubles en la saliva por medio de algunas de sus cualidades químicas y papilas gustativas.
¢ son las encargadas de captar y distinguir los sabores.

8.1 La lengua y los receptores gustativos
Los órganos del Gusto, que tienen por misión el percibir y enviar al cerebro el sabor de las cosas que introducimos en la boca, se encuentran en los Bulbos o botones gustativos, localizados en la Lengua.
Lengua (anatomía), órgano musculoso de la boca, asiento principal del gusto y parte importante en la fonación y en la masticación y deglución de los alimentos. La lengua está cubierta por una membrana mucosa, y se extiende desde el hueso hioides en la parte posterior de la boca hacia los labios. La cara superior, los lados y la parte anterior de la cara inferior son libres. El resto está unido a la cavidad bucal. Los músculos extrínsecos fijan la lengua a distintos puntos externos y los músculos intrínsecos, que discurren de forma vertical, transversal y longitudinal, permiten muchos y diversos movimientos.
Es ésta un órgano musculoso fijo por la base al suelo de la boca y con la punta libre, de forma que puede realizar toda clase de movimientos. La superficie de la lengua está cubierta por una mucosa que tiene una serie de salientes denominados Papilas Linguales que son de diferentes formas, las bases de estás papilas tienen numerosas terminaciones nerviosas. Cuando una sustancia penetra en la boca es disuelta por la saliva produciendo una corriente nerviosa que nos produce la sensación del gusto, la cual es transmitida al cerebro a través de los nervios correspondientes. La lengua tiene otras utilidades como es ayudar en la masticación e ingestión de los alimentos, y sobretodo en la articulación de las palabras cuando hablamos (las consonantes principalmente).

1 . Los receptores sensoriales y los órganos de los sentidos. Los receptores sensoriales son estructuras que contienen células especializadas en detectar determinados tipos de variaciones del medio ambiente, cuando estas variaciones superan un determinado valor (umbral) originan un impulso nervioso que se transmiten a través de las neuronas. Estos tipos de variaciones reciben el nombre de "estímulos". Los receptores sensoriales pueden estar dispersos por el cuerpo, como pasa con los receptores sensoriales de temperatura, o pueden estar agrupados constituyendo los denominados "órganos de los sentidos", como los que constituyen los ojos o el oído.
2 . Tipo de receptores sensoriales. Los receptores sensoriales se clasifican según el tipo de estímulo que captan en:
- Mecanorreceptores (captan efectos mecánicos), como los receptores del tacto de la piel (sentido del tacto), los del equilibrio del oído interno y los de la audición del caracol del oído (sentido del oído).
- Termorreceptores (captan temperaturas) como los termorreceptores de la piel.
- Quimiorreceptores (captan sustancias químicas) como las mucosas olfativas de la nariz (sentido del olfato) y las papilas gustativas de la lengua (sentido del gusto).
- Fotorreceptores (captan luz) como la retina del ojo (sentido de la vista).
Los quimiorreceptores de la lengua. Son los responsables del sentido del gusto, es decir de la captación de las sustancias disueltas en los líquidos. Son las células sensibles que forman los botones gustativos que se encuentran en unas protuberancias de la lengua denominadas papilas
8.2 Transducción ácida
Está causado por ácidos, y la intensidad de la sensación gustativa es aproximadamente proporcional a la concentración de iones hidrógeno. En otras palabras, cuanto más fuerte es el ácido, más intensa la sensación.
Al igual que el sabor salado, se produce por compuestos ionizables, pero donde juega un papel predominante los iones hidrógeno (hidrogenión) como cationes, por ejemplo: HCl, H2SO4, CH3COOH, etc. En este casos el hidrogenión bloquea directamente a los canales de K+ de la membrana de las células gustativas, inhibiendo la salida de iones K+. Este aumento de cargas
positiva aumenta el potencial de membrana, activando a los canales de Ca++ dependientes de voltaje.
Debido a que el mecanismo es parecido al de los sabores salados, algunas sales muy fuertes pueden inclusive tener un sabor agrio.
La intensidad del sabor agrio depende de dos factores: primero, al igual que los compuestos salados, la capacidad de ionizarse que tenga el compuesto debido al tamaño del anión; y segundo, el pH que tenga el compuesto (mientras más ácido es, más agrio sabe). Estos dos motivos explican porque el HCl es más agrio que CH3COOH.
Este sabor se detecta con más facilidad en las regiones posterolaterales del dorso de la lengua, territorio básicamente perteneciente a las papilas foliadas.

8.3 Transducción salada
El gusto salado depende de sales ionizadas. La calidad del gusto varía algo de una sal a otra, porque las sales también estimulan otros botones gustativos en grado variable.
Si a un poco de agua le agregamos un poco de sal de cocina, percibimos una sensación particular, sobre todo en los bordes laterales de la lengua, que es acompañado por una secreción fugaz de saliva.
Se relaciona más a los compuestos inorgánicos debido a la facilidad con que estos se ionizan en soluciones, entre ellos los cloruros: cloruro de sodio, de potasio, calcio, litio, etc; y los hálidos: hálido de bromuro, yodo, fluoruro, etc. en el caso de las sustancias saladas, el catión (ion positivo) produce el cambio en la célula, pero según el tamaño del anión es la intensidad del sabor. Esto se debe a que mientras más pequeño es el anión, más disociable es el compuesto (por el tipo de unión química) y más fácilmente habrá interacción entre el catión y la célula. Así, el NaCl es más salado que el acetato de Na+ (CH3COONa).
Al disociarse el compuesto, el catión (por ejemplo, el Na+) entra por canales iónicos y, al haber entrada de iones positivos, el potencial de membrana de la célula se positiviza, activando a los canales de Ca++ dependientes de voltaje.
Este sabor se percibe más fuertemente en las regiones anterolaterales del dorso de la lengua, territorio también de las papilas fungiformes.

8.4 Transducción amarga
Al igual que las sustancias dulces se caracterizan por ser compuestos orgánicos pero dentro de este
gran grupo hay dos grupos más específicos para los sabores amargos: los alcaloides, encontrados en
muchos medicamentos y otras sustancias como: cafeína, nicotina, estricnina, quinina, etc; y sustancias
orgánicas de cadena larga, especialmente si contienen grupos nitrógeno.
Esta similitud entre las moléculas dulces y amargas hace que algunas sustancias que saben dulce de
inicio luego dejen un sabor amargo (por ejemplo la sacarina y el alcohol).
Estas sustancias se unen a receptores unidos a una proteína Gq (no estimulatoria sino regulatorio), la cual activa una enzima fosfolipasa. Esta enzima desdobla el PIP2 en DAG y de más significancia en IP3.
Este IP3 se une a canales de Ca++ dependientes de IP3 en el retículo endoplásmico de las célula gustativa, aumentando la liberación masiva de iones Ca++ al espacio intracelular. Luego de aquí ya se inicia la vía común.
El sabor amargo se percibe más intensamente cerca de las papilas circunvaladas y en el tercio
posterior de la lengua.
· Notarán que estos dos últimos sabores: el agrio y el amargo, se perciben fuertemente en áreas posteriores de la lengua. Ya que la mayoría de las sustancias tóxicas producen sabores amargos (por ejemplo la estricnina) esto permite un mecanismo de defensa para el individuo pues se produce un sabor “desagradable” antes de que la sustancia pueda ser deglutida.

8.5 Transducción dulce
No depende de ninguna clase aislada de productos químicos. Una lista de algunos productos químicos que causan este sabor es la siguiente: azúcares, glicoles, alcoholes, aldehídos, cetonas, amidas, ésteres, aminoácidos, etc. Obsérvese específicamente que casi todas las sustancias que causan sabor dulce son productos químicos orgánicos.
Aparte de las sustancias obvias como la glucosa, sacarosa y demás carbohidratos, hay otros compuestos orgánicos que producen un sabor dulce, como: glicéridos, alcohol, aminoácidos, cloroformo, cetonas, amidas y ésteres, entre otras.
Cuando la sustancia dulce se pone en contacto con el botón gustativo, estimula un receptor acoplado a una proteína Gs (G estimulatorio). Esta proteína Gs luego estimula una enzima adenilciclasa, la cual aumenta la formación de AMPc intracelular. El AMPc, a través de una PKA (proteínkinasa A) bloquea los canales de K+. Al no poder salir K+ de la célula, por la carga positiva, el potencial de membrana va aumentando hasta activar los canales de Ca++ dependientes de voltaje, siguiendo la vía común.
Otra vía por la que la sustancia dulce positiviza el potencial de membrana de la célula gustativa es a través de la excitación de unos canales de Na+ de amilorida. Estos canales se abren y permiten la entrada de iones de carga positiva (a pesar de su nombre no son totalmente específicos para el Na+)
lo cual activará a los canales de Ca++ y a partir de ahí sigue la vía común.
El sabor dulce se percibe con más intensidad en la punta de la lengua, territorio básicamente de las papilas fungiformes.

BIBLIOGRAFIA
http://www.monografias.com/trabajos14/elgusto/elgusto.shtml
http://www.aula2005.com/html/cn3eso/13organssentits/13organssentitses.htm
http://www.pucmmsti.edu.do/cienciasfisiologicas/FIS-LNB1.PDF

TEMA 7. VISION.
TEMA 7. VISION
la visión, es la facultad por la cual a través de un órgano visual, se percibe el
entorno que rodea al sujeto. Muchos organismos vivos poseen receptores fotosensibles capaces de reaccionar ante determinados movimientos y estímulos luminosos, pero la verdadera visión supone la formación de imágenes en el cerebro. Los ojos de los distintos organismos perciben imágenes de diversas características, generalmente diferentes a las que percibimos los seres humanos.
7.1 LUZ VISIBLE
La luz visible es una de las formas como se desplaza la energía. Las ondas de luz son el resultado de vibraciones de campos eléctricos y magnéticos, y es por esto que son una forma de radiación electromagnética (EM). La luz visible es tan sólo uno de los muchos tipos de radiación EM, y ocupa un pequeño rango de la totalidad del espectro electromagnético. Sin embargo, podemos percibir la luz directamente con nuestros ojos, y por la gran importancia que tiene para nosotros, elevamos la importancia de esta pequeña ventana en el espectro de rayos EM.
Las ondas de luz tienen longitudes de onda entre 400 y 700 nanómetros (4 000 y 7 000 Å). A medida que el arcoiris se llena de matices, nuestros ojos perciben diferentes longitudes de ondas de luz. La luz roja tiene longitudes de onda relativamente largas, aproximadamente 700 nm (10-9 metros) de largo. La luz azul y la luz morada tienen ondas cortas, aproximadamente 400 nm. Las ondas más cortas vibran a mayores frecuencias, y tienen energías más elevadas. Las luz roja tiene una frecuencia aproximada de 430 terahertz, mientras que la frecuencia de la luz azul es de aproximadamente 750 terahertz. Los fotones rojos tienen aproximadamente 1.8 electrón-Volt(eV) de energía, mientras que cada fotón azul transmite aproximadamente 3.1 eV.
Los vecinos de la luz visible en el espectro EM son la radiación infrarroja de un lado, y luz ultravioleta del otro lado. La radiación infrarroja tiene longitudes de ondas más largas que la luz roja, es por esto que oscila a una frecuencia menor y lleva consigo menor energía. La radiación ultravioleta tiene longitudes de ondas más cortas que la luz azul o violeta, por lo que oscila más rápidamente, y porta mayor cantidad de energía por protón que la luz visible.
La luz viaja a la increíble velocidad de 299 792 458 kilometros por segundo (aproximadamente 186 282.4 millas por segundo). A esta increíble velocidad, ¡la luz podría girar más de siete veces alrededor de la Tierra en cada segundo!. La letra "c" minúscula se usa en las ecuaciones para representar la velocidad de la luz, como es el caso de la famosa relación entre energía y materia de Einstein: "E = mc2". Todas las formas de ondas electromagnéticas, incluyendo los rayos X y las ondas de radio , y todas las demás frecuencias a lo largo del espectro EM, también viajan a la velocidad de la luz. La luz viaja más rapidamente en el vacío, y se mueve más lentamente en materiales como agua o vídrio
7.2 EL OJO Y LAS CÉLULAS FOTORECEPTORAS
Los ojos son unos órganos complejos que se desarrollan básicamente como ventanas laterales del cerebro. Cada ojo presenta una capa de receptores, un sistema de lentes para enfocar la imagen sobre estos receptores y un conjunto de terminaciones nerviosas encargadas de transmitir los potenciales de acción hacia el cerebro. El ojo está constituido por dos segmentos de esfera. El segmento mayor (también de radio mayor) está en la parte posterior y se manifiesta externamente como la esclerótica o "blanco del ojo". En la parte anterior, el segmento pequeño de la esfera (de menor radio) se denomina córnea, se trata de una cubierta externa transparente, con una capa epitelial escamosa estratificada y transparente y cuya superficie exterior está bañada por una fina película de lágrimas; su función es lubricante, desinfectante y de limpieza (Fig. 1).
 FIG. 1
FIG. 1El espacio existente detrás del cristalino, órgano con forma de lente biconvexa, contiene un material gelatinoso, el humor vítreo, cuyo ritmo de recambio es muy lento. En la porción periférica del cristalino se extiende el cuerpo ciliar; que contiene los procesos ciliares, por los que la coroides se une al cristalino y al músculo ciliar. Las cámaras anterior y posterior (situadas por delante y por detrás del iris) están llenas de humor acuoso, líquido transparente parecido al LCR, segregado activamente por los procesos ciliares. Su velocidad de producción y absorción es lo suficientemente elevada para reemplazar el volumen total de la cámara varias veces durante el día. El humor acuoso fluye desde la cámara posterior a la anterior a través de la pupila y después se absorbe hacia el sistema venoso en el ángulo que existe entre la córnea y el iris (por un sistema de trabéculas y canales). El desequilibrio entre la secreción y reabsorción de humor acuoso puede hacer aumentar la presión intraocular, lo que pone en peligro la viabilidad de la retina. Esta alteración patológica es conocida como glaucoma.
El reflejo de acomodación del cristalino (Fig. 3) consiste en una modificación del poder refractario de la lente lo que posibilita que se forme una imagen enfocada en la retina. Mientras que los peces, anfibios y serpientes acomodan desplazando el cristalino y modificando su distancia a la retina, los restantes reptiles, aves y mamíferos no alteran la posición de la lente, sino que el mecanismo de acomodación consiste en la modificación activa de la curvatura del cristalino por contracción de los músculos ciliares.
Si el cristalino fuera extraido del ojo, tomaría una forma esférica debido a la forma esférica de la capsula que lo rodea. En el ojo en reposo (visión lejana), los ligamentos suspensorios tiran sobre el ecuador del cristalino causando un aplanamiento en su eje anteroposterior. Este aplanamiento hace menos convexo al cristalino y provoca una menor refracción de la luz lo que permite que en la retina se puedan enfocar objetos a más de 6 m de distancia (ojo en reposo u ojo acomodado).
Para enfocar imágenes de objetos cercanos al ojo, el cristalino debe tomar una forma convexa más esférica. Esto se produce por la contracción de los músculos ciliares. El resultado de este proceso es que el cristalino se hace más esférico con una mayor capacidad de refracción de la luz lo que permite que se enfoquen en la retina objetos más cercanos (ojo enfocado u ojo no acomodado). Estos mecanismos de acomodación requieren un globo ocular resistente que evite la deformación de la retina, y un humor vítreo muy viscoso que impida retrodesplazamientos del cristalino. El mecanismo de acomodación del cristalino es un reflejo parasimpático que se inicia por percepción de imágenes borrosas procedentes de la retina.
 FIG.2
FIG.2La capacidad de acomodación del cristalino disminuye con la edad, debido a que éste se hace menos elástico y no puede incrementar su curvatura como se requiere para la visión cercana. Este trastorno aparece con frecuencia en la especie humana a partir de los 40 años de edad y se denomina presbiopía o vista cansada.
Reflejo de apertura pupilar (Fig. 3). El iris es una membrana pigmentada con forma de disco, perforada centralmente, que forma una cortina para regular la cantidad de luz que entra al ojo. La pupila es el orificio situado aproximadamente en el centro del iris. El iris contiene fibras musculares lisas capaces de modificar el diámetro de la pupila y consiguientemente, la cantidad de luz que penetra en el ojo. El músculo dilatador pupilar contiene fibras radiales inervadas por nervios simpáticos del ganglio cervical craneal; la contracción de estas fibras produce midriasis (aumento del diámetro pupilar). El músculo constrictor pupilar (esfínter pupilar) contiene fibras circulares con gran capacidad de acortamiento inervadas por fibras parasimpáticas del nervio oculomotor común y su contracción produce miosis (disminución del diámetro pupilar).
En el ojo de los animales vertebrados las imágenes se proyectan sobre la retina a través de la córnea y las lentes (pupila y cristalino) formando una imagen más pequeña, invertida y de dos dimensiones. El área visual de la corteza cerebral será la responsable de formar una imagen igual a la real: a tamaño natural, derecha, en tres dimensiones y coloreada (según especie) (Ver Fig. 1).
La luz, una vez que atraviesa la córnea, el iris y el cristalino, es absorbida por los pigmentos visuales de las células receptoras de la retina, codificando las imágenes ópticas en impulsos nerviosos, cuya actividad eléctrica varía con la cantidad de luz recibida.
El desarrollo de la retina no es paralelo al desarrollo evolutivo de los vertebrados. Por ej., la retina de los primates es más simple anatómica y funcionalmente que la retina de una rana o una paloma, aunque exista una relación directa entre la complejidad de la retina de un animal y el desarrollo de los centros visuales de su cerebro. En nuestros animales domésticos la retina y corteza visual más desarrollada es la del gato.
 FIG. 3
FIG. 3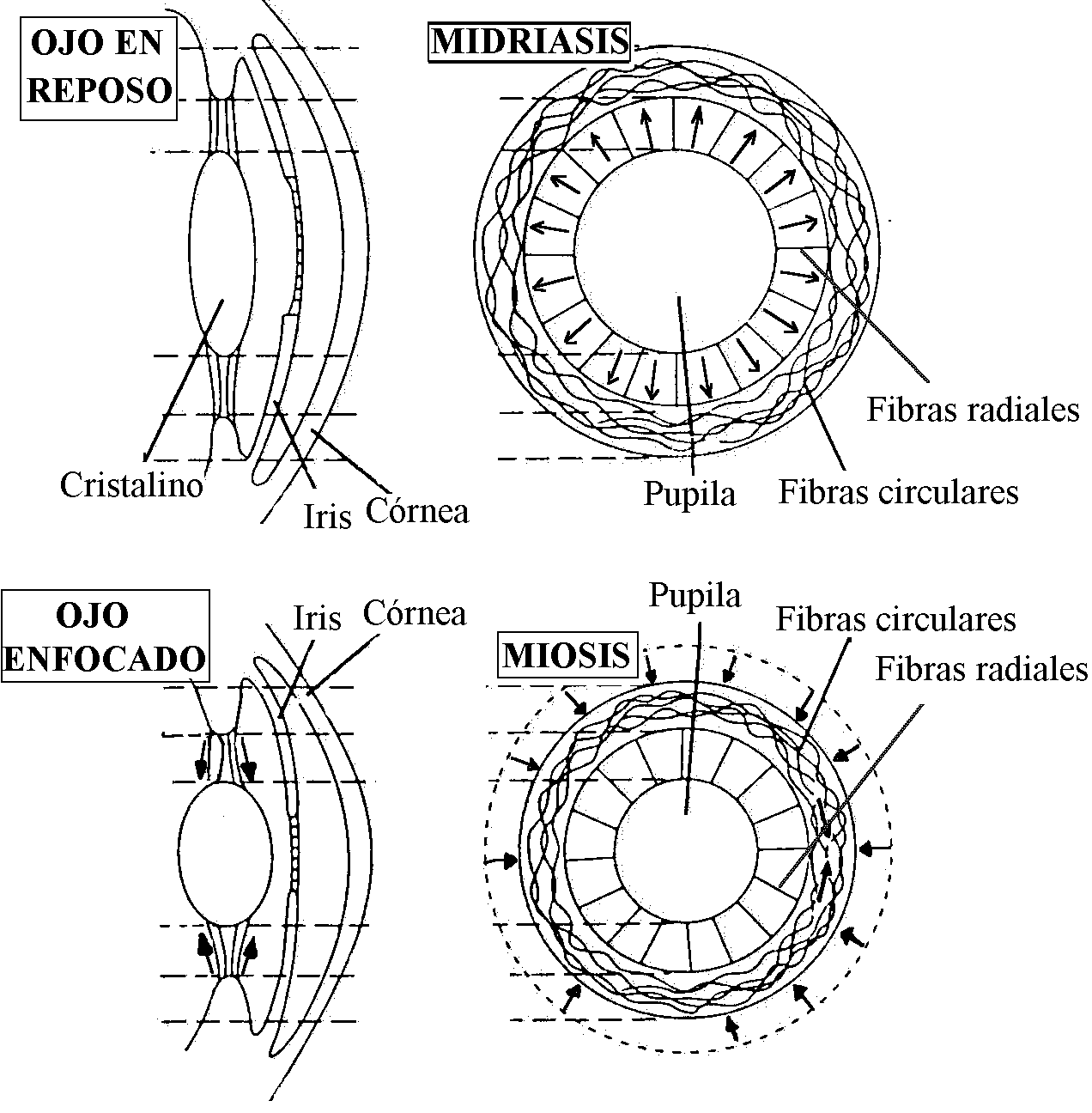 FIG. 4
FIG. 4Existen dos tipos principales de células fotorreceptoras (bastones y conos). Ambos tipos celulares establecen una conexión sináptica directa con interneuroras, células bipolares, que conectan a las células fotorreceptoras con las células ganglionares. Los axones de estas últimas llevan los potenciales de acción hacia el cerebro a través del nervio óptico. Modificando el flujo de información en las sinapsis entre fotorreceptores, células bipolares y células ganglionares existen dos tipos celulares de interneuronas: las células horizontales y las amacrinas. Las primeras están entre los fotorreceptores y las células bipolares y establecen sinapsis de inhibición lateral para conseguir una visión más perfecta; mientras que las amacrinas se disponen mediando entre las células bipolares y las ganglionares.
 FIG.5
FIG.5En un gran número de animales (la mayoría de los animales domésticos y silvestres pero no en el cerdo y hombre) esta capa pigmentaria contiene un pigmento reflectante en la zona denominada tapetum (tapetum lucidum), de tal manera que la luz que llega al tapetum se refleja y regresa de nuevo a los fotorreceptores lo que permite que la retina puede hacer un uso óptimo de la luz que llega a los ojos en condiciones de menos intensidad luminosa, aunque a expensas de la agudeza visual; el tapetum es el responsable del brillo en los ojos de los animales cuando se les dirige una luz en la oscuridad.
A lo largo de la mayor parte de la retina, los rayos de luz atraviesan las 10 capas retinianas y las células ganglionares, bipolares y ocasionalmente amacrinas y horizontales antes de alcanzar los fotorreceptores. Aunque, membranas, núcleos y células son relativamente transparentes pueden, sin embargo, provocar alguna distorsión de los rayos de luz. La fóvea (de 0,4 mm de Ø) situada en el centro de la retina es la zona de la retina donde se logra la mayor agudeza visual. Está especialmente diseñada para minimizar la distorsión de la luz, ya que en ella, las células ganglionares y bipolares están colocadas hacia un lado permitiendo el acceso directo de los rayos luminosos a los *conos, pues los bastones se encuentran fuera de la fóvea (Fig. 5).
En la exploración ocular, utilizando un oftalmoscopio se puede observar un punto donde la retina se halla interrumpida por la salida de los axones de la capa celular ganglionar en su camino hacia el cerebro. Este punto, localizado en posición anterior a la fóvea, es el disco óptico; origen del nervio óptico (I par craneal), nervio muy rico en axones (existen más axones en ambos nervios ópticos que en todas las raíces dorsales de la médula espinal). En el disco óptico no existen fotorreceptores, por lo que también se denomina área ciega. También visible con el oftalmoscopio es la salida de los vasos sanguíneos retinales. Éstos son una trama de arterias y venas que penetran a la retina a nivel del disco óptico y proveen gran parte de la nutrición de la retina; los vasos de la coroides proporcionan la nutrición restante.
Los fotorreceptores, de los cuales existen 120 millones de conos y 7 millones de bastones en la especie humana, tienen características morfológicas similares, pero son diferentes funcionalmente. En ambos tipos celulares se distinguen tres regiones comunes claramente diferenciadas: el segmento externo, el segmento interno y el terminal sináptico (Fig. 6).
El segmento externo está especializado en la fotorrecepción y presenta una elaborada ordenación de unos 1.000 discos membranosos apilados ricos en pigmentos visuales (rodopsina en el bastón y yodopsina en el cono); los discos en los bastones están separados de la membrana plasmática al contrario de lo que ocurre en los conos. El segmento interno conecta con el externo mediante un cilio modificado que contiene nueve pares de microtúbulos, pero que carece de los dos pares de microtúbulos centrales; el segmento interno contiene el núcleo, las mitocondrias y demás estructuras citoplasmáticas. El terminal sináptico, por último, entra en contacto con una o más células bipolares.
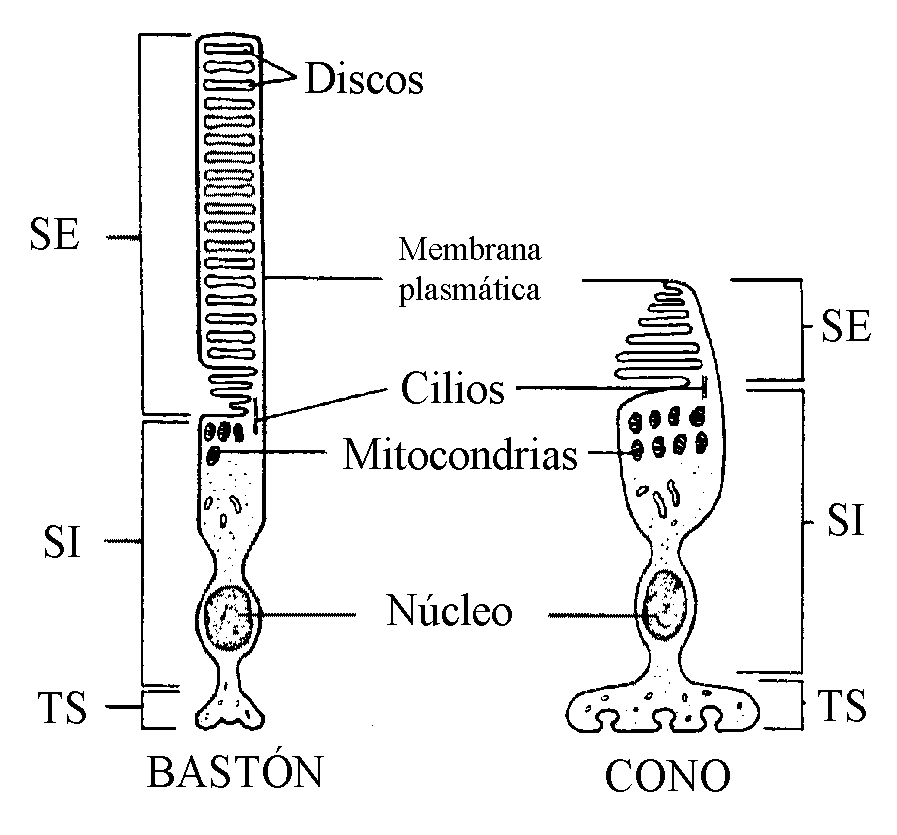 FIG.6
FIG.67.3 FOTOQUÍMICA DE LA VISIÓN
En los fotorreceptores se produce una transducción foto-quimio-eléctrica que da lugar a que en la terminal sináptica se libere mayor o menor cantidad de NT en relación con la magnitud del potencial receptor.
La estimulación de los fotorreceptores se inicia por la absorción de la luz por el pigmento visual y el efecto fotoquímico correspondiente; ello lleva consigo cambios de permeabilidad iónica y génesis de potencial receptor que, desde el segmento externo, pasa al segmento interno y se transmite a la región sináptica donde, mediada por un transmisor, la señal alcanza otras neuronas retinianas (bipolares y horizontales).
Los fotorreceptores son distintos al resto de receptores sensoriales pues no detectan impulsos nerviosos típicos; sin embargo, en los bastones y conos, al ser alcanzados por la luz, se establecen unas ciertas condiciones físico-químicas que van a desencadenar el impulso de otras células nerviosas. Donde si se detectan los impulsos nerviosos es en el nervio óptico, por fibras nerviosas de las células ganglionares.
Sin lugar a equivocarnos, la fotoquímica de la visión es el mecanismo más complicado y preciso de los sentidos. Este complejo mecanismo lo estudiamos en tres apartados: ciclo del pigmento visual, generación del potencial receptor y adaptación a la luz y oscuridad.
1) CICLO DEL PIGMENTO VISUAL. Los pigmentos visuales son proteínas complejas; pero se ha visto que la parte del pigmento que absorbe la luz (porción cromatófora) es una sustancia muy parecida a la vitamina A, se trata del aldehído de la vitamina A (retinal) en sus formas cis y trans. Los pigmentos visuales de la membrana fotosensible de bastones y conos son diferentes. En los bastones se encuentra la rodopsina y en los conos hay yodopsina. Para explicar los mecanismos fotoquímicos de la visión nos referiremos a los bastones por estar mejor estudiados (Fig. 7).
 FIG.7
FIG.7La rodopsina se sintetiza en ausencia relativa de luz y su síntesis implica la actuación de una enzima con aporte de energía metabólica para la reducción de todo el trans-retinal a cis-retinal. Después, este cis-retinal se recombina con la opsina para formar de nuevo rodopsina.
2) GENERACIÓN DEL POTENCIAL RECEPTOR. En los conos y bastones no existe potencial de acción, sólo un potencial receptor que se transmite al resto de las células nerviosas, siendo las células ganglionares las encargadas de transmitir los potenciales de acción a través del nervio óptico.
El mecanismo de producción del potencial receptor (Fig. 7) es el siguiente: la bomba Na+/K+ está restringida a la membrana que rodea el núcleo y el segmento interno impulsa continuamente iones Na+ desde el interior al exterior y, por tanto, crea un potencial negativo dentro de la célula. Sin embargo, en oscuridad, la membrana del segmento externo se hace permeable y deja pasar fácilmente el Na+ y así neutraliza en gran parte la negatividad del interior de toda la célula, dando un potencial receptor a los bastones de -25 a -30 mV. Este potencial receptor es proporcional al logaritmo de la intensidad de la luz, y así el ojo puede distinguir entre intensidades luminosas muy variadas. Cuando la rodopsina se expone a la luz, se descompone y esto hace disminuir la conductancia de los iones Na+ hacia el interior del bastón, aunque sigan impulsándose iones desde el segmento interno hacia el exterior. De esta forma, resulta que hay salida de iones positivos sin la correspondiente entrada de los mismos por el segmento externo; lo que produce aumento de la negatividad intracelular (estado de hiperpolarización), alcanzándose valores de -90 mV.
Puesto que la membrana plasmática del bastón está separada de la de los discos que contienen el pigmento fotosensible, el efecto de la disminución de la permeabilidad para el Na+ debe depende de un mediador químico como es el GMPc. Al parecer, el GMPc se encarga de mantener los canales de Na+ en configuración abierta; la luz activa una proteína, llamada transducina, en la membrana del fotorreceptor que promueve la acción de una fosfodiesterasa que hidroliza el GMPc, lo que hace que los canales de Na+ se cierren y la membrana se hiperpolarize.
3) Adaptación a la luz y a la oscuridad. Los ojos son capaces de adaptarse a niveles altos y bajos de intensidad luminosa. La adaptación a la luz ocurre cuando el animal es expuesto a la luz brillante, como al salir de un establo en un día soleado. Esto provoca que las sustancias fotoquímicas de los bastones y conos se reduzcan a opsinas y retinal, lo que hace disminuir la sensibilidad del ojo a la luz. Al mismo tiempo, el diámetro de la pupila se reduce por constricción refleja parasimpática del músculo constrictor pupilar, disminuyendo la cantidad de luz que entra al ojo.
La adaptación a la oscuridad ocurre cuando el animal se desplaza de un ambiente bien iluminado a un lugar oscuro, o más gradualmente al oscurecer en la tarde. En la oscuridad, todo el retinal está incorporado a la rodopsina y gran parte de la vitamina A del epitelio pigmentario es absorbida por los bastones cuyo contenido en rodopsina será máximo y su sensibilidad a la luz también. De esta forma, la reconstitución de las sustancias fotoquímicas permite a los ojos detectar niveles muy bajos de intensidad luminosa.
7.4 VISIÓN A COLOR
La esfera ocular no es ninguna maravilla de la ingeniería. Es sólamente una estructura que aloja la retina y le proporciona imágenes enfocadas y nítidas del mundo exterior. La luz entra en el ojo a través de la córnea y el iris, atravesándo la lente del cristalino antes del alcanzar la retina.

La retina recibe una pequeña imagen invertida de ese mundo exterior, transmitida por el sistema óptico formado por la córnea y el cristalino. El ojo es así una pequeña 'cámara oscura'. La lente del cristalino altera su forma para enfocar la imagen, pero esa capacidad adaptativa se va perdiendo con la edad, por lo que perdemos capacidad visual óptica.

El ojo es capaz de adaptarse a distintos niveles de iluminación gracias a que el diafragma formado por el iris puede cambiar de diámetro, proporcionando un agujero central (la pupila) que varía entre 2 mm (para iluminación intensa) y 8 mm (para situaciones de poca iluminación).
La retina traduce la señal luminosa en señales nerviosas. Está formada por tres capas de células nerviosas. Sorprendentemente, las células fotosensibles (conocidas como conos(cones) y bastones(rods)) forman la pate trasera de la retina (es decir: La más alejada de la apertura del ojo). Por eso, la luz debe atravesar antes las otras dos capas de células para estimular los conos y los bastones.

Las causas e historia evolutiva de este diseño invertido de la retina no se conocen bien, pero es posible que esa posición de las células fotosensibles en la zona más posterior de la retina permita que cualquier señal luminosa dispersa sea absorbida por las células pigmentarias situadas inmediatamente detrás de la retina, ya que contienen un pigmento oscuro conocido como melanina.
Puede también que estas células con melanina ayuden a restaurar químicamente el equilibrio del pigmento fotosensible de los conos y bastones cuando éste pierde su capacidad debido al desgaste causado por la acción de la luz.
La capa media de la retina contiene tres tipos de células nerviosas: Bipolares, horizontales y amacrinas. La conexión de los conos y bastones con estos tres conjuntos de células es complejo, pero las señales terminan por llegar a la zona frontal de la retina, para abandonar el ojo a través del nervio óptico. Este diseño inverso de la retina hace que el nervio óptico tenga que atravesarla, lo que da como resultado el llamado punto ciego (blind spot) o disco óptico.
Los bastones y conos contienen pigmentos visuales, que son como los demás pigmentos en el sentido de que absorben la luz dependiendo de la longitud de onda de ésta. Sin embargo, estos pigmentos visuales tienen la particularidad de que cuando un pigmento absorbe un fotón de energía luminosa, la forma molecular cambia y se libera energía.
El pigmento que ha cambiado su estructura absorbe peor la energía y por eso se dice que se ha blanqueado o despigmentado (bleached). La liberación de energía por parte del pigmento y el cambio en la forma molecular hacen que la célula libere una señal eléctrica mediante un mecanismo que aun no se conoce por completo.
Los bastones son sensibles a niveles muy bajos de iluminación y son los responsables de nuestra capacidad de ver con poca luz (visión escotópica). Contienen un pigmento cuyo máximo de sensibilidad se halla en la zona de los 510 nanómetros (o sea, la zona de los verdes). Al pigmento de los bastones, la rodopsina, se la suele llamar 'púrpura visual', ya que cuando los químicos logran extraerlo en cantidad suficiente, tienen una apariencia púrpurea.
La visión escotópica carece de color, ya que una función de sensibilidad con un espéctro único es ajena al color, por lo que la visión escotópica es monocromática.
Los conos son los que proporcionan la visión en color. Hay tres clases de conos. Cada una de ellos contiene un pigmento fotosensible distinto. Los tres pigmentos tienen su capacidad máxima de absorción hacia los 430, 530 y 560 nanómetros de longitud de onda, respectivamente. Por eso se los suele llamar "azules", "verdes" y "rojos". No es que los conos se llamen así por su pigmentación, sino por el supuesto 'color de la luz' al que tienen una sensibilidad óptima.

Esta terminología es bastante desafortunada, ya que las luces monócromas de 430, 530 y 560 nm. de longitud de onda no causan realmente la percepción de azul, verde y rojo, sino la de violeta, azul verdoso y amarillo verdoso. Por eso, las denominaciones conos cortos, conos medios y conos largos (por el tipo de longitud de onda al que son sensibles comparativamente) es más lógica (las abreviaciones en inglés son: S-cones (cortos), M-cones (medios) y L-cones (largos)).
La existencia de tres funciones de sensibilidad espectral proporciona la base de la visión en color, ya que cada longitud de onda causará una proporción única de respuestas en los conos sensibles a longitudes cortas, medias y largas. Son los conos quienes nos proporcionan la visión en color (visión fotópica), que permite distinguir notablemente bien pequeños cambios en la composición de longitudes de onda de una luz.
7.5 Percepción
La percepción del color depende tanto de las longitudes de onda de luz que refleja la superficie del objeto, como de la composición espectral de la fuente de iluminación, que puede variar considerablemente en el tiempo y de un lugar a otro, del área circundante al objeto y del estado de adaptación del observador. La capacidad del sistema visual para compensar todas estas variaciones, de modo que los objetos parezcan tener el mismo color, se conoce como constancia del color.
Percepción visual
La percepción visual es el complejo proceso de recepción e interpretación significativa de cualquier información recibida. Ojo y cerebro tienden a comprender y organizar lo que vemos imponiéndole un sentido racional aunque particularizado por la experiencia de cada individuo. Tras esa primera función de reconocimiento, nuestro sentido de la percepción entra en una fase analítica que comprende la interpretación y organización del estímulo percibido, mediante la cual se estructuran los elementos de esa información, distinguiendo entre fondo y figura, contornos, tamaños, contrastes, colores, grupos, etc. Igualmente por la percepción tendemos a complementar aquellos elementos que puedan dar definición, simetría, continuidad, unificación y "buena forma" a la información visual.
La decodificación del significado de la información visual va a depender también de otros factores de influencia recopilados por la experiencia personal e intelectual de cada indivíduo, lo que en definitiva hacen que la misma se realice bajo un aspecto puramente subjetivo.
La decodificación del significado de la información visual va a depender también de otros factores de influencia recopilados por la experiencia personal e intelectual de cada indivíduo, lo que en definitiva hacen que la misma se realice bajo un aspecto puramente subjetivo.
PERCEPCIÓN VISUAL Proceso cognoscitivo de recepción e interpretación de la información recibida mediante un estímulo visual IMAGEN como representación visual de una forma o de una idea | |
OJO y CEREBRO Comprenden y Organizan la información contenida en el estímulo visual | |
Fases de la Percepción | |
1. Reconocimiento | 2. Análisis |
Aceptación y comprensión de la información recibida | Interpretación y Organización racional del estímulo |
Experiencia intelectual del receptor | |
Subjetiva. Selectiva. Temporal. | |
Niveles de la Percepción | |
Instintivo. Descriptivo. Simbólico | |
PROCESO DE LA PERCEPCION VISUAL
- El objeto físico emite o refleja radiaciones luminosas de distinta frecuencia e intensidad (estímulos).
- Las radiaciones luminosas penetran en el interior del globo ocular a través de la pupila, que se dilata o contrae en función de las condiciones luminosas por la acción del iris. Hasta llegar a la retina, que es la parte fotosensible del ojo, tiene que atravesar la córnea, el cristalino y la cámara interior acuosa.
- La retina está compuesta por tres tipos de células (ganglionares, bipolares y fotorreceptoras). La energía luminosa incide primero sobre las células ganglionares que no son sensibles a la luz; luego sobre las bipolares que tampoco lo son y, por último, sobre las fotorreceptoras (conos y bastoncillos) que son las únicas sensibles a la luz. Hay, además, otros dos tipos de células: las células horizontales, que conectan entre sí a los conos y los bastoncillos, y las células amacrinas, que conectan las células bipolares con las ganglionares.
- Sólo una pequeña parte del espectro electromagnético de las ondas luminosas (el situado entre las 380 y las 780 milimicras) puede ser captada por los conos y bastoncillos, los cuales están especializados. Los conos actúan como receptores del color y operan en condiciones de moderada o alta iluminación ambiental. Los bastoncillos son receptores acromáticos que operan en condiciones de escasa iluminación ambiental. En el ojo humano hay unos 120 millones de bastoncillos y entre 6 y 7 millones de conos. En total, pues, hay más de 126 millones de células fotorreceptoras.
- En estas condiciones, los estímulos luminosos producen en la retina del observador una proyección óptica invertida del objeto. El tamaño de la proyección óptica varía según sea la distancia entre el objeto y el observador. La forma de la proyección óptica varía con el cambio de la inclinación del objeto respecto al observador.
- La energía electromagnética que incide sobre los conos y bastoncillos es transformada en impulsos nerviosos que llegan hasta las células ganglionares cuyos axones se unen para formar el nervio óptico en el disco óptico, llamado punto ciego porque carece de células fotorreceptoras y no es sensible a la luz.
- Los haces nerviosos de cada ojo se encuentran en el quiasma óptico donde parte de ellos se cruzan para ir a parar al hemisferio cerebral opuesto, de manera que las fibras que salen del lado izquierdo de ambas retinas (y que corresponden al lado derecho del campo visual) se proyectan hacia el hemisferio izquierdo y las que salen del lado derecho de ambas retinas (y que corresponden al lado izquierdo del campo visual) se proyectan hacia el hemisferio derecho.
- Los impulsos nerviosos llegan a través del cuerpo geniculado lateral del tálamo hasta la corteza visual del cerebro, situada en el lóbulo occipital, modifican su estado fisiológico y se produce la experiencia perceptiva

BIBLIOGRAFIA
http://centros5.pntic.mec.es/ies.arquitecto.peridis/percep/pvisual2.html
http://www.uco.es/organiza/departamentos/publicaciones/fisiovet/tema4.html
http://www.visionyvida.org/lavision/visionDelColor.htm
http://www.aloj.us.es/galba/DIGITAL/CUATRIMESTRE_II/IMAGEN-PAGINA/1codigos1.htm
http://www.gusgsm.com/funciona_ojo_humano
Suscribirse a:
Comentarios (Atom)